

In climatology, global warming or climate change indicates the alteration in the earth’s climate that developed from the end of the 19th century to the beginning of the 20th century and is still ongoing [1]; it is generally represented by inflation in the global average temperature and associated atmospheric phenomena (e.g., drought [2], heatwaves [3], increase in extreme phenomena linked to the water cycle such as floods, desertification [4], melting ice, rising ocean levels [5], and changes to atmospheric circulation patterns with storms, hurricanes, and phenomena that are more intense such as cyclones [6]). On one hand, the anthropical increase in the amount of greenhouse gases (GHGs) such as ozone (O3), water vapor (H2O), methane (CH4), carbon dioxide (CO2), chlorofluorocarbons (CFCs), and nitrous oxide (N2O) in the atmosphere and, on the other, the natural variation in the incident solar radiation due to the Milankovitch cycle, have led to these dangerous phenomena [7,8,9]. By the end of the 21st century, the global temperatures under different scenarios (i.e., global climate models characterized by the representative concentration pathways and a set of alternative plausible trajectories of future global development (SSPs)) are projected to increment by +1.18 °C/100 years. (SSP1 2.6), +3.22 °C/100 yearrs. (SSP2 4.5), +5.50 °C/100 years. (SSP3 7.0), and +7.20 °C/100 years. (SSP5 8.5), with greater warming oriented over the Northern hemisphere’s high latitudes and feebler warming over the Southern hemisphere [10].
The steadfast propensity to global warming has had a severe impact on overall viticulture [11]. Given the observed increase in growing season temperatures in Europe (+1.7 °C from 1950 to 2004), between 2000 and 2049, it is assumed that temperatures for major wine regions will increase on average by about +0.42 °C per decade and will generally increase by +2.04 °C [12,13]. Districts usually entertained as highly desirable for grapevine growing can now show an excess of heat load when confronted with the needs of the currently grown selective breeding [14]. In these areas, global warming could randomize the appropriateness of wine growing and synchronously lead to a switch in grape cultivars’ cultivation, creating extensive damage to wineries subject to production regulations [15].
The typical microclimate of the area is fundamental for characterizing the particular style of wine produced in very specific areas [16]. Obtaining the complete and balanced ripening of the bunch is essential for determining the best time for harvesting and this is highly influenced by climatic variability which poses new challenges for winemakers from year to year [17]. Several models have predicted that all assessed grape phenological phases will undergo anticipation that will be significant for the véraison and the maturity stages (until 15 days before) with respect to the flowering phase [18]. All this can translate into a series of problems in the berry, such as the presence of unbalanced qualitative parameters [19]. In fact, the worsening of drought can lead to an excess of the sugar content of the berry and a 40% reduction in production [20]. Owing to a loss in membrane stability of Oenococcus oeni cells, caused by high alcohol content, malolactic fermentation can negatively be triggered (i.e., undesirable sensory alteration and lag in wine stabilization) [21,22]. In addition, an excess of total soluble solids leads to the formation of undesirable fermentation by-products (e.g., acetic acid and glycerol) with up-regulating glycolytic and pentose phosphate pathway genes [23].
Phenolic compound development is affected by genetic factors as well as environmental parameters such as ultraviolet (UV) radiation, sunlight, maximum and minimum temperatures, and grapevine water status [24,25,26]. The berry composition (i.e., total soluble solids, proanthocyanidins, flavan-3-ol monomers, and pigmented polymers; [27]) is impacted by cultivation practices (soil management and canopy handling strategies) and the existing microclimate around the developing vines [28,29,30].
This review, after a summary of the synthesis of phenolic compounds in the berry (flavonoids and non-flavonoids), describes the impact of climate change and therefore of environmental elements (such as temperature, water, solar radiation, and UV-B light) on the berries accumulation and storage throughout ripening and harvesting. The soil and canopy management techniques achieved in the field to mitigate the degradation of phenolic compounds and preserve their concentration are also further discussed.
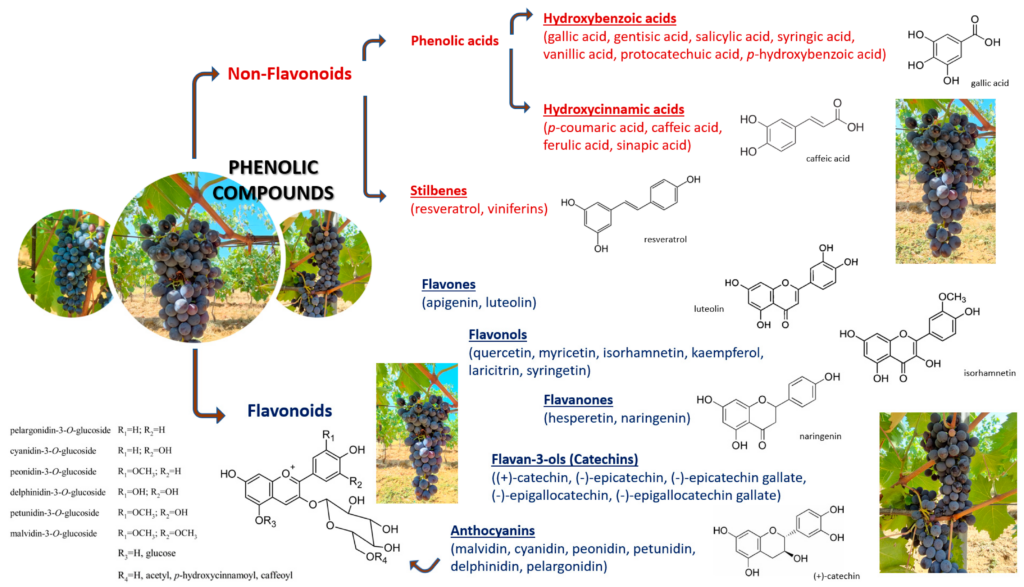
Figure 1. Classification of phenolic compounds
Global warming alludes to long-term variations in the climate of Earth affected by warmth imbalances from natural origin (e.g., volcanic, solar, or internal dynamics) and anthropogenic sources (e.g., greenhouse gas ejections and soil management) [116,117,118]. Below is an image to highlight and underline how the global temperature is undergoing a change (Figure 3).
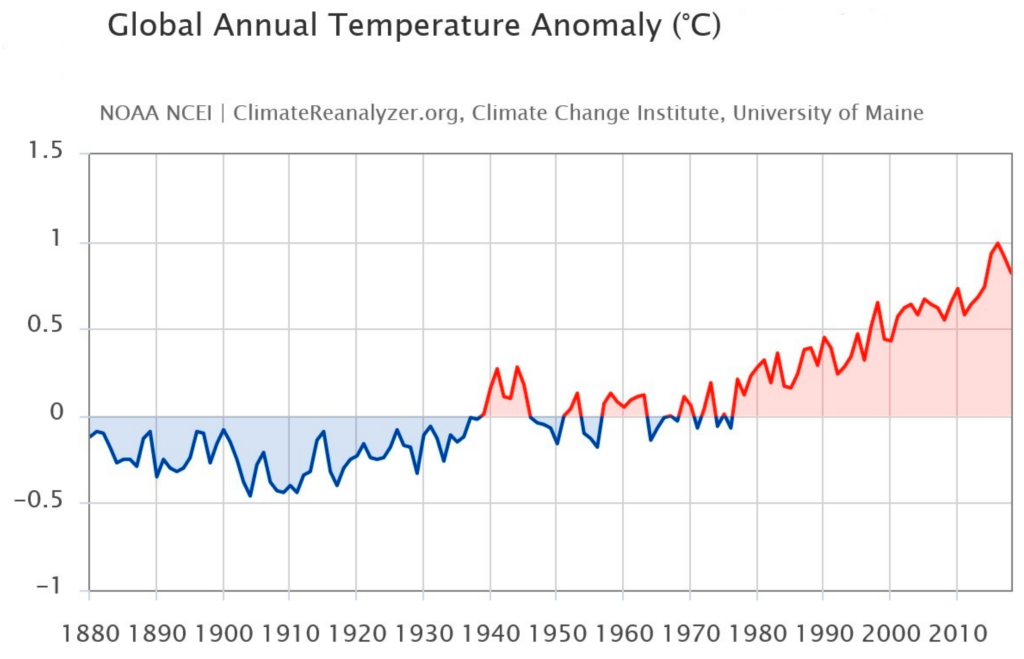
Figure 3. Images from Birkel, S.D. ‘Explore Climate Change’, Climate Reanalyzer (https://ClimateReanalyzer.org), Climate Change Institute, University of Maine, USA. Accessed on 22 June 2023. Global annual temperature anomaly (°C) 1901–2000 baseline [119].
In addition, Figure 4 supplies a selection of template maps and time-series performing modifications in temperature and precipitation over the past century and how they are predicted to shift in the future (RPC 8.5) [120].
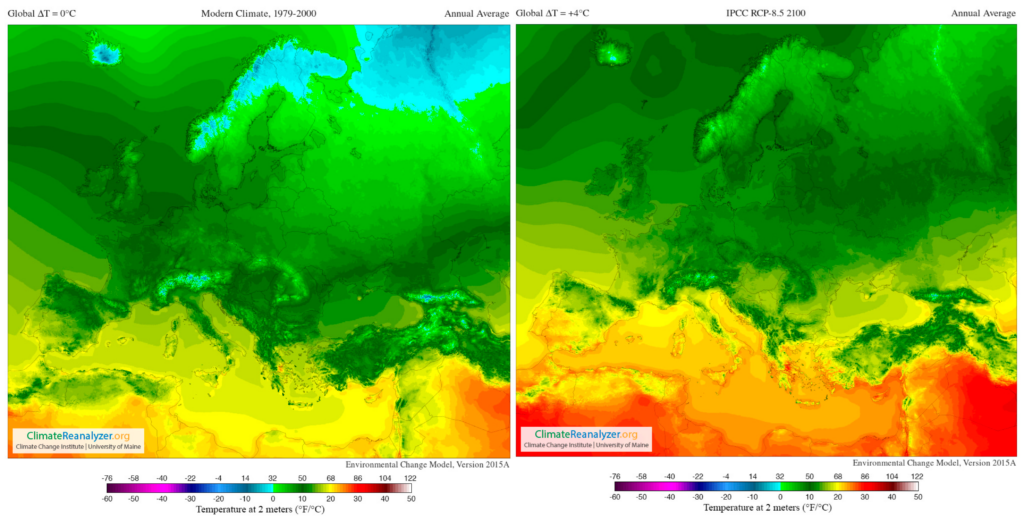
Figure 4. Images from Birkel, S.D. ‘Environmental Change Model’, Climate Reanalyzer (https://ClimateReanalyzer.org), Climate Change Institute, University of Maine, USA. Accessed on 22 June 2023. In the Environmental Change Model (ECM), solutions are calculated from gridded inputs of monthly temperature and precipitation using a degree day solver and biome rubric. Boundary conditions for a given experiment are derived by blending reanalysis (modern climate) and general circulation model (future climate) climatologies. The ratio of reanalysis to the future climate depends on a user-selected global temperature departure value, ∆T. For the modern climate, ∆T = 0 °C; for 2100, ∆T = +4 °C (European model) [119].
The most important viticultural regions in Europe are situated equatorward top at the 37–50° N parallels [121]; as seen in the figures, these areas are significantly affected by climate change (present and future). In the short-term, the climate markedly governs the whole ripening development process of grapes, because it demands appropriate temperatures, radiation intensities, as well as specific standards of water availability throughout the growth and maturation cycle, ultimately influencing berry characteristics [121,122].
Water
During the last decennium, in a lot of viticultural areas, pressure on water resources increased owing to global warming being associated with drought conditions. So, the interest in water use optimization to guarantee crop sustainability is urging researchers to introduce new approaches for reducing water losses (e.g., partial root-zone drying and regulated deficit irrigation) [123].
A recent RNA-sequencing analysis work carried out on white grapes reported that water deficit can affect the berries’ composition [124]. In particular, chalcone synthases, two chalcone isomerases, one flavonoid-3′5′-hydroxylase, two flavanone-3-hydroxylases, one dihydroflavonol reductase were up-regulated by deficit irrigation (−1.5 MPa ΨStem of vines at 67 days after anthesis). Water deficit significantly promoted the expression of flavonol synthase (a key enzyme for flavonol production). Leucoanthocyanidin reductase (VviLAR1) and anthocyanidin reductase (as well-known regulators of the flavan-3-ols and proanthocyanidin biosynthesis) were also up-regulated by water deficit.
Anthocyanins’ hydroxylation and methoxylation degrees in berries were altered by environmental markers, such as the water status [125]. With partial rootzone drying (PRD) management, a relative reduction in the proportion of methoxylated anthocyanins in grape berries was found [126], owing to the enhancement in endogenous abscisic acid levels [127]. Dihydroxylated and non-acylated forms have high antioxidant capacity, whereas trihydroxylated and methoxylated are the most oxidized anthocyanins [128]. Shellie and Bowen [129] noted increases in monomeric anthocyanins under water deficit, but possibly by an indirect effect such as a reduction in the size of the berry; (i.e., skin-to-pulp ratio increased). Increments by water deficit in the relative abundance of methoxylated anthocyanins were also found (most oxidized forms) [125]. In this experiment, the procyanidin epicatechin gallate, and the flavonols quercetin and quercetin-3-glucoside declined; however, total low molecular weight phenols were not significantly reduced by water deficit. Several authors observed an increase in total anthocyanins in grapes of plants subjected to water stress [130,131]. However, it was recently observed that an excess of water stress (associated with heatwaves; 35 °C) led to the reduction in extractable anthocyanins in the must and to an increase in quercetin-3-O-glucoside and quercetin-3-O-glucuronide [132,133]. An interesting experiment showed two types of berry responses to drought stress: (I) indirect and positive action on phenolic compounds concentration owing to berry size reduction; (II) direct effect on biosynthesis that can be positive or negative depending on the phenolic compound type, application period, and severity of drought [134]. Briefly, flavonols’ biosynthesis was higher for medium water stress (early water deficit between anthesis and véraison) and strong water stress (between véraison and harvest maturity); biosynthesis of flavan-3-ols was reduced by the early water deficit between anthesis and véraison; and biosynthesis of proanthocyanins and anthocyanins increased only for late significant water deficit between véraison and harvest maturity. In each case, the drought stress enhanced the degree of tannin polymerization.
Berries skin with an intermediate deficit (40% of field capacity) showed a higher content of trisubstituted anthocyanins [135]. Structural genes of the phenylpropanoid pathway which encode flavonoid 3′ (F3′H) and 3′,5′-hydroxylases (F3′5′H) explain the biosynthesis of cyanidin-based and delphinidin-based anthocyanin pigments. The alternative accumulation of trisubstituted rather than disubstituted anthocyanins is attributed to a fork in the pathway and is joined to the competitive activity of F3′H and F3′5′H [136]. The elevated amount of trisubstituted anthocyanins (major color stability in red wines [57]) promotes greater resistance to oxidative damage [137]. Guidoni et al. [138] found that 3′-substituted anthocyanin biosynthesis was probably more strongly influenced by environmental conditions (and cultural practices) than was 3′,5′-substituted ones. In fact, the peonidin-3-glucoside/malvidin-3-glucoside ratio differed on average from 1.9 during the hot and rainy season to 1.2 in the cooler and very dry season. Regulated deficit irrigation (RDI) normally refers to all irrigation strategies that take vines at some degree of water deficit for a prescribed part of the season [139]. According to these results, in Tempranillo grapevines, a step-by-step upgrade in water deficit as summer progressed (ideally from −0.2 to −0.9 MPa) (that implied a greater water deficit between véraison and harvest when anthocyanin synthesis happens) encouraged the synthesis of malvidin (i.e., the main tri-hydroxylated anthocyanin), compared with water stress caused in the berry herbaceous stage growth (RTD_1), or even with RDI_2 that included an additional stress period shortly after véraison. On the contrary, RDI water management resulted in a higher proportion of non-acylated delphinidine and petunidine. In contrast, some results showed that an altered grapevine water status would impact the skin flavonoids’ biosynthesis only slightly, and its effect would not depend that much on the vine water status, but on the seasons [140,141]; additionally, in a recent work, no evidence that the water stress level caused significant differences in monomeric flavan-3-ol compounds was found [142].
A recent work [135] showed the pre-véraison water deficit effects on vines grafted on Mgt 101-14 and 1103 Paulsen rootstocks. From the obtained data, the presence of VIT_18s0001g03430 (VvFLS gene; it encodes a flavonol synthase) transcript was enhanced in severe deficit (25% of field capacity). A study [143] where irrigation was applied every week (when Ψpd reached −0.4 MPa; moderate water deficit) indicated no significant key metabolites modified (phenolic acids, stilbenoid DP1 and DP2, flavonols, flavan-3-ols, and di-OH anthocyanins and tri-OH anthocyanins) by treatments.
Considering the presence of contrasting works regarding the effects of water deficit alone, a more in-depth investigation into potted plants (controlled environment) with multiple cultivars and multiple rootstocks to which different levels of stress can be applied without other variables would be desirable. Quantitative, metabolomic, and transcriptomic analyzes during bunch ripening could provide answers to doubts regarding water scarcity and its repercussions on quality.
Temperature
The Intergovernmental Panel on Climate Change has denoted the ineluctable increase in worldwide temperature and has recognized climate change as an important menace to the overall food supply [144]. Environmental temperature is a climate element that influences phenological phases and causes the timing of ripening and harvest [145].
In general, an increased temperature in the plant will increase its rate of metabolic processes with an associated rise in metabolite accumulation [146]. However, the following statement would need to be corrected from a climate change perspective for the reasons discussed below. Merlot berries’ extreme temperatures exposure for short periods in ripening (blew forced air across the bunches; to a target temperature 2 × ∆Tr degrees above the average; where ∆Tr represented the temperature difference between the sun bunches average and the shade bunches average) altered the anthocyanins’ partitioning between acylated and non-acylated and between dihydroxylated and trihydroxylated forms in the biosynthetic pathway. Briefly in this trial, with ten combinations, the exposure hours varied from 2 to 200 and the arbitrary thresholds were >30 °C, >35 °C, and >40 °C. Higher berry temperatures led to a higher concentration of malvidin-based anthocyanins, driven chiefly by growth in the acylated derivatives (i.e., malvidin 3-coumaroyl-glucoside). At berry temperatures tantamount to those of shaded ones, exposure to solar radiation reduced the acylated forms of the five base anthocyanins and augmented the proportion of dihydroxylated anthocyanins [147]. Other authors found that temperature did not influence the accumulation of malvidin 3-glucoside in Merlot [148] or in Pinot noir [149].
qRT-PCR analysis proved that temperature (and light conditions) affected the anthocyanin composition in the skin through the regulation of the flavonoid biosynthesis pathway genes (VlMYBA1-3, VlMYBA1-2, and VlMYBA2) [150]. These results suggested that low temperature (and light) had a synergistic incidence on gene expression in the flavonoid biosynthesis pathway; high temperature (>34 °C) (or dark treatment) severely suppressed anthocyanin accumulation. The authors affirmed that the skin anthocyanin composition in the 35 °C (and dark) treatment could not be defined (the content was too low). In 15 °C (and light), 40.2% peonidin and 28.1% malvidin derivatives were found; in addition, malvidin derivatives’ levels decreased (about 17.5%) in 15 °C (and dark), and peonidin (30.7%) and malvidin (7.3%) derivatives decreased in 35 °C (and light) treatment.
Peonidin-3-monoglucoside was the prevailing anthocyanin form in 15 °C (and light) treatment, but its percentage dwindled in two treatments: 15 °C (and dark; 5.1%) and 35 °C (and light; 4.8%). Moreover, the peonidin-3-p-coumarylglucoside-5-glucoside augmented in 15 °C (and dark; 24.0%) and 35 °C (and light; 14.0%) treatments. Moreover, chemical composition changes were also correlated with warmer seasons, as indicated by the rising formation of malvidin, petunidin, and delphinidin coumaroyl derivatives [146].
In sunlit growth chambers (rotating and stationary phytotron rooms), optimal requirements for anthocyanin accumulation (Cardinal, Pinot noir, and Tokay grapevines) came about when bunches were set out to cool nights (15 °C) and temperate days (25 °C) during maturation; this deep coloration was not found in clusters ripened at 35 °C day combined with 15 °C night, 35 °C day combined with 25 °C night, and 15 °C day combined with 25 °C night temperatures. Moreover, a day temperature of 35 °C entirely obstructed anthocyanin synthesis in Tokay grapes, regardless of night temperatures [151]. In agreement with the results just stated, high temperatures (35 °C) cut down the total anthocyanin content (Cabernet Sauvignon cultivar) with respect to the control berries (25 °C), with the exception of malvidin-3-glucoside, malvidin-3-acetylglucoside, and malvidin-3-p-coumaroylglucoside. This reduction resulted from factors such as anthocyanin degradation and from the inhibition of mRNA transcription of the anthocyanin biosynthetic genes (qRT-PCR and LC-MS analysis of 13C-labelled anthocyanin were carried out) [152].
The temperature effects on proanthocyanidin biosynthesis are understood to a small degree. A recent study on Cabernet Sauvignon grape berries cultured in vitro showed that the proanthocyanidin content was higher in berries (skin + seeds) cultured at a low temperature compared with a high one. In addition, the elevated temperatures forbade the expression level of anthocyanidin reductase and leucoanthocyanidin reductase-1 (key genes involved in proanthocyanidin biosynthesis; ANR and LAR-1) [153].
The consequence of temperature on tannins is not yet well known; one study on Merlot berries showed that high temperature during berry development heightened the start of skin tannin storage, however this effect was not evident at véraison [154]. Meanwhile, another trial on high temperature during berry ripening did not survey any interaction on skin tannins by vintage in Sangiovese [155]. Maybe, as stated by Gouot et al. [156], a deficiency of knowledge of the accurate galloylation process limits gene expression investigations to genes involved in the phenylpropanoid pathway. It was hypothesized that temperature could affect tannin biosynthesis and galloylation in young green berries, owing to modifications in the regulation of transcripts coding for glucose-gallic acid-glucosyltransferase [19].
In a recent study, a rise in galloylated flavan-3-ol subunits (berries skin) and galloylation percentage skin tannins was noted two weeks after the end of the treatment (heated bunches max 45 °C) [157].
Considering these results, in view of climate change, it would be interesting to monitor the evolution of the phenolic component during heatwaves (before, during, and immediately after the phenomenon). Furthermore, more studies would be desirable to understand the changes in the hydroxycinnamic compounds (there are still scant studies in this regard).
Solar Radiation and UV-B Light
Sunlight is chiefly comprised of infrared wavelengths but also contains 8–9% ultraviolet light (UV). UV light can be subdivided into three types: UV-A (315 to 400 nm; not absorbed by the O3 layer); UV-B (280 to 315 nm; its intensity on Earth’s surface depends on the O3 layer thickness); and UV-C (100 to 280 nm; completely absorbed by the O3 layer) [158].
Shaded fruit showed lower flavonol glucosides at vintage and during berry development in many cultivars such as Cabernet Sauvignon, Shiraz [159], and Merlot [146,160]. In fact, it was suggested [161] that the flavonol profile is a reliable indicator to estimate canopy architecture and grapes’ exposure to solar radiation. Blancquaert et al. [162] suggested that UV-B radiation plays a notable role in the photo-protection of the berry against light exposure. In their experiment, the light quantity and quality were, respectively, manipulated in the bunch zone with leaf removal and by installing ultraviolet B-suppression sheets within the bunch zone. Moreover, treatments showed only a marginal effect on skin flavan-3-ol synthesis and no impact on seed tannin. In relation to the hydroxycinnamic acid, hydroxybenzoic acid, and stilbene content, no significant correlations were found with ambient sunlight intensity [158]. So, these authors suggested that in the ripening of berries, flavonols operate as UV screening compounds, defending plant tissue from light damage (i.e., higher flavonol glucosides). In fact, light influences flavonol synthase (VvFLS) expression, a significant flavonol structural gene [163], and influences a transcriptional regulator of flavonoid synthesis (VvMYBF1) [164]. Nevertheless, UV-B is also known to up-regulate genes encoding phenylalanine ammonia-lyase and naringenin–chalcone synthase [165]. At BBCH 73 stage (groat-sized berries), responsiveness to UV increased, as detected by the induction of class III chitinase, phenylalanine ammonia-lyase, and stilbene synthase expression, together with accumulation of resveratrol [166]. Flavan-3-ols and proanthocyanidins in berry skins were affected by sun exposure. In fact, as demonstrated by [163], shadow consistently decreased the molar ratio of the tri-hydroxylated gallocatechins to di-hydroxylated catechins and decreased the mean degree of polymerization. Meanwhile, light exposure increased the proanthocyanidins galloylation rate in berries after the removal of shading.
As concerns the results of skin anthocyanin, data highlighted a complex, combined effect of solar radiation + berry temperature; in particular, there is a synergistic effect with quite moderate berry temperatures and an antagonistic effect at elevated temperature extremes. An association of low light summarized to high berry temperature dropped the total concentrations of skin anthocyanin. Instead, quercetin 3-glucoside (flavonol-glycoside) enhanced with exposure to solar radiation [147]. These results were confirmed by other authors [132,133]; high quantities were found of quercetin-3-O-glucoside and quercetin-3-O-glucuronide in stressed grapes. Vijay and Vadewki [167], in their experiment on Cabernet Sauvignon grapes, explained the benefit of uncovered clusters to solar light; in treated clusters (leaf removal, cluster thinning, and shoot thinning) higher concentrations of flavan-3-ols (i.e., catechin and epicatechin) and flavonols (i.e., quercetin and myricetin) were found (quercetin accumulation could be a light solar-dependent process). Another recent study adopting high UV-B radiation (Vitroflex 395 filter and UV-B lamps) on Tempranillo grapes showed an increase in quercetin and kaempferol contents correlated to the expression of flavonol synthase and chalcone synthase genes (VvFLS4 and VvCHS1), and an increase in hydroxycinnamic acids at the pea-size stage [168]. On the contrary, in UV-exposed grape skins, hydroxybenzoic acids showed no significant alterations [169]. In the group of non-flavonoid phenolic acids, Singleton et al. [170] hypothesized that the trans-configuration of coutaric and caftaric acids was a natural phenomenon and the cis-form was the UV-induced isomerization product.
In light of all these interesting works, it is deemed necessary to further investigate the effects of the interaction between light radiation and ambient temperature.
Another possible consideration is to carry out in-depth studies on the effects of solar radiation on the bunch, focused on the individual cultivars and on the organoleptic effects on the wines. In fact, in the light of recent studies [171], increases in the must of quercetin (Sangiovese variety) could lead to the impoverishment of the finished product (precipitated by the hydrolysis of glycosides; i.e., by the supersaturation of aglycones) [172].
Management Tips: How Can the Winegrower Corroborate the Grapevine Resilience?
The strategies discussed below are integrated into a respectful and conservative approach to the vineyard ecosystem. These soil or canopy managements are included in the narrative as they are considered natural resources that can increase the resilience of the plant with a view to preserving the finished product for the consumer. The use of these substances defends the plant from abiotic stress and their application is compliant with organic agriculture. The arbitrariness of use means that they can be applied preventatively and occasionally only during critical years [173]. Furthermore, the emphasis to be placed on these products is the fact of corroborating the system and not distorting the plant’s eco-physiological functions to arrive at a “standardized product”, but rather of supporting and creating adaptation to abiotic stress.
Zeolite
Zeolites are porous aluminosilicates characterized by a crystalline frame that comprehends an interconnected chambers/channels system. Their geometrical parameters are notable characteristics responsible for their adsorption skills. Consequently, zeolites can function as a sorbent for environmental pollutants, a water reservoir (reversible dehydration), and nutrients for plants (cations + anions) [174] (Figure 5).
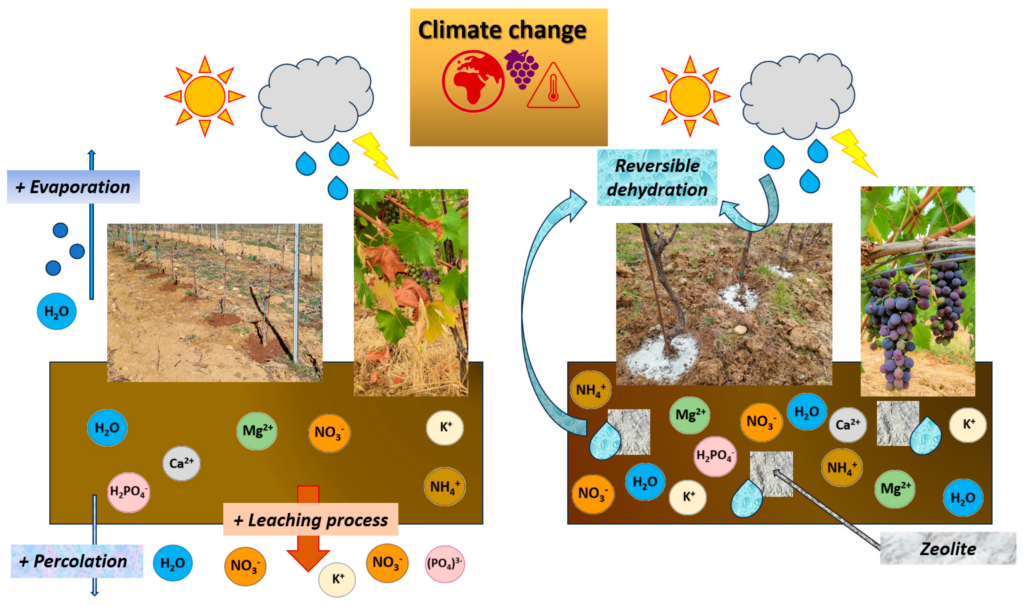
Figure 5. Effects of zeolite on vineyard soil that mitigate the effects of climate change (adapted from [174]).
Owing to their rare properties, these aluminosilicates have become more and more trendy in recent years and found functional applications in viticulture [175]. Among natural zeolites, clinoptilolite represents the most commonly applied in agriculture and environmental defenses [176].
In a recent study [177], it was seen that canopy treatments of chabasite-rich zeolite (15 kg/L ha−1) increased the content of phenolic compounds in the grapes, as well as providing simultaneous control of gray mold (Botrytis cinerea; Botryotinia fuckeliana) and Lobesia botrana (Denis and Schiffermüller). This probably happened owing to its ability to be an inert mineral able to reduce canopy surface temperatures as well as being radiation-reflecting. Other authors found an increase in extractable anthocyanins together with a reduction in quercetin forms in plants treated with soil applications of clinoptilolite [132,133]. This could be attributable to the ability of zeolites to mitigate water stress and allow for balanced ripening in extremely dry years. However, there are still too few studies on the use of this ductile mineral on grapevines; only recently has research been turning to its possible uses to tackle climate change. Given its abundance in nature and given its ease of application, this corroborant product could represent a valid vineyard management strategy in critical years.
5.2. Kaolin
Al2Si2O5(OH)4 (i.e., kaolin) is a mineral rock consisting predominantly of white kaolinite that is inert, harmless, and is skillfully squanders in water. Kaolin’s force, once sprayed, is correlated to the white safeguarding particle film that gathers on the grapevine leaf’s surface; this particle film is able to augment excess radiation reflection, by reducing heat load accumulation damage and UV solar detriments [178].
As shown in Figure 6, it was found that kaolin enhanced the total monomeric anthocyanins amount in Malbec [179] and Meili berries [180], reducing their sensibility to temperature and solar exposure.

A recent study confirmed that kaolin treatments induced an increase in total phenols (+13.0%), flavonoids (+10.5%), tannins (+27.5%), and ortho-diphenols (+30.0%) confronted with the control treatment [183]. Bernardo et al. [184] confirmed that ortho-diphenols, flavonoids, and tannins were ameliorated in kaolin-treated plants.
All of these experiments highlighted the salient role of weather conditions in triggering the accumulation of phenolic processes. Moreover, they showed that kaolin-treated vines exhibited a major response to oxidative stress markers by enhancing secondary metabolite storage in hot harvests. The employment of kaolin supported different cultivars reinforcing their performance in alleviating severe drought, high temperature, and sunburned stress impacts.
Seaweed Extract
Biostimulant products are recognized as formulated biological substances that ameliorate the productivity of plants as a result of the novel or emergent properties of the complex of constituents, plant growth regulators, and/or plant safeguarding complexes; they are auxiliary tools to improve vine nutrition and to develop vine resilience to environmental stress [185].
Ascophyllum nodosum can be a sustainable management tool to lessen the short-period shock of severe temperatures and strong light irradiance in grapevines; in fact, it was found that it was able to encourage a prompt photosynthetic recovery after drought stress [184]. In addition to this, numerous studies have highlighted its positive effect on the phenolic composition of berries [189,192,194].
Conclusions
It is believed that techniques to allay the effects of climate change on grapevine clusters and phenolic compounds should be taken from a “single case” perspective. It is not possible to standardize and approve treatments or agronomic techniques for each different case in different environments (for example, different cultivars, different legislations, and obligations). In order to intensify the resilience of the plant and therefore to maintain or implement its performance in terms of secondary metabolites, it is necessary to evaluate the “single vineyard case”. In fact, by connecting to the concept of terroir (which can be defined as that well-defined area where the natural, physical, and chemical conditions, the geographical area, and the climate allow the achievement of a fixed wine) it is necessary today to establish an elastic and variable approach towards the single wine year, moving away from the concept of product standardization. In fact, a different approach and characterization of the finished wine by the winegrowers and the consumers are desirable, or rather a tolerant point of view of a product susceptible to change based on the vintage (color, aromatic intensity, tannic persistence, etc.). Considering the product in its entirety (climatic changes and adaptations) and therefore in its specific peculiarities of the single vintage could lead to an evolution of techniques and approaches towards these fascinating ecosystems. On the other hand, becoming stuck in the spasmodic search for maintaining the same organoleptic characteristics of wine (different vintages) could lead to a loss of innovation and a loss of new organoleptic perceptions.